
by Wesley Dáttilo, Thiago Junqueira Izzo and Víctor Rico-Gray
Many plant species, mostly in the tropics, have extrafloral nectaries (EFN-bearing plants) that usually secrete a nutritious liquid. This liquid attracts ants that, in exchange for food, protect their host plants against potential herbivores. Within a natural environment different ant and plant species can interact with each other and generate complex interaction networks. In these networks, ant and plant species are represented as objects called nodes and their interactions are represented by links (Figure 1). In our recent studies we have used tools derived from graph theory to investigate the organization of these ant-plant networks in different ecosystems around the world.
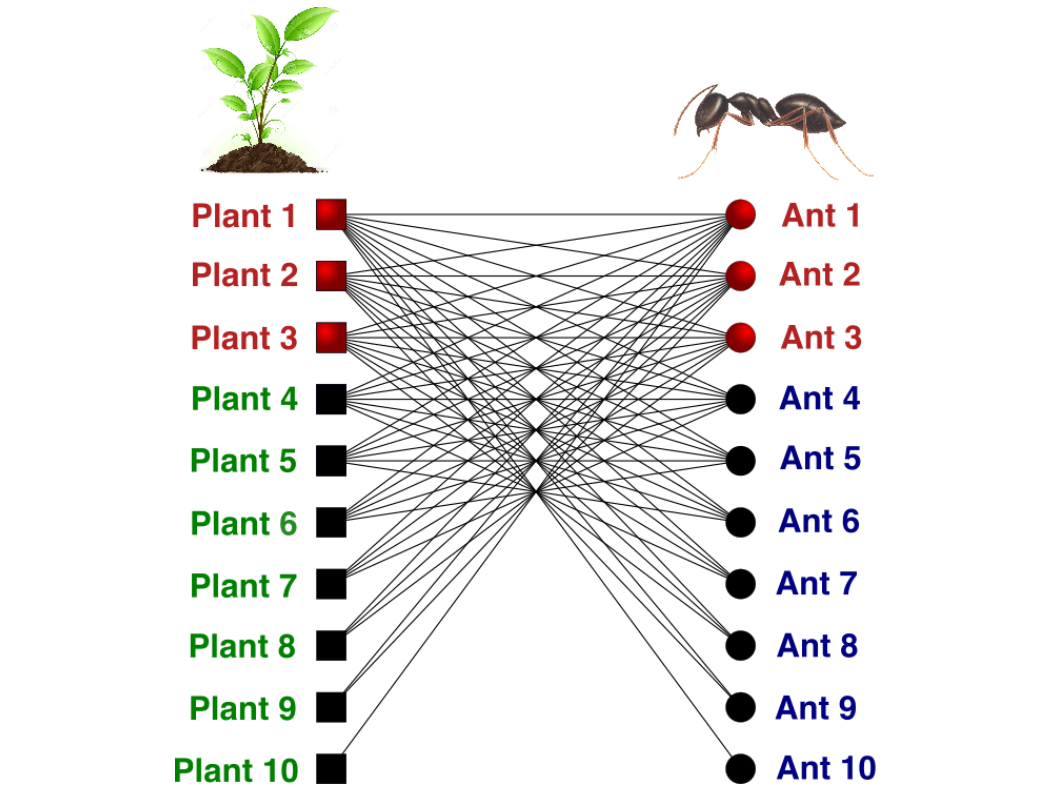
Figure 1. Ecological network involving EFN-bearing plants (squares) and ants (circles). Each node represents one plant or ant species, and lines represent ant–plant interactions. Plants and ants with red nodes are those species present in the generalist core of the network, and all other species represent the periphery of the network. This network is ordered according to the nested pattern (see text for more information).
One of the main characteristics of ant-plant interaction networks involving EFN-bearing plants is that there is a central core of highly interacting species formed by both ants and plants that interact more strongly with each other compared to other peripheral species (those with fewer interactions). Once this central core is virtually connected with all species in an environment, these species have the capacity to influence the ecological and evolutionary dynamics of the whole system. However, what are the factors that could generate differences in the number of interactions among species within an environment? In other words: why are some species associated to the generalist core and others to the periphery of the networks?
Several factors have been proposed to explain the origin of structural patterns in ecological networks. A key factor that describes the heterogeneity of interactions in a system is based only on the difference in the relative species abundance (neutral models). In this case, abundant species should interact most frequently with each other and with less abundant species, forming a core of highly connected species. In contrast, less abundant species will rarely interact among themselves.
Studying ants associated to EFN-bearing plants in the Brazilian Amazon we (Dáttilo et al. 2014a) show that the central core of ants and plants interact with each other more than expected by neutral models based only on the species abundance in an environment. In addition, central ant species within the generalist core were those with eco-physiological adaptations to feed on liquid food, mainly species of the genera Azteca, Brachymyrmex, Camponotus, Crematogaster, and Dolichoderus. In another study (Dáttilo et al. 2013), we also demonstrated that even though the ant and plant composition of networks changed over space, the central core of generalist species and the network structure remained unaltered on a geographic distance of up to 5 km. This finding indicates that independent of variation in local and landscape environmental factors, the nonrandom pattern organization of these interacting assemblages does not change and all coevolutionary pressure exerted by the core species are stable at a regional scale.
Additionally, other studies published by the same research group show that the central core of highly interacting ant species is stable over long time scales or even after the perturbations generated by tropical hurricanes. So, why is this central core of ant species so stable? Motivated by this question we studied the interactions between ants and EFN-bearing plants in a tropical coastal environment on the Gulf of Mexico (Dáttilo et al. 2014b). In this study, we investigated the role of ant dominance hierarchy in structuring the nested pattern of ant-plant. This pattern of interaction predicts that interactions are organized such that specialist ants interact with subsets of the plant species with whom generalist ants interact. (Figure 1). We found that within a nested ant–plant network, ant species found in the central core of highly interacting species were competitively superior, showing massive recruitment and resource domination, compared with peripheral species with fewer interactions (Figure 2). Since the number of ants on the host plant is associated with effectiveness in defense against herbivores, it is possible that most plant species found within ant–plant networks could be better protected against herbivory by these dominant ant species. In short, these results highlight the importance of competition and monopolization in the resource use by ants in the maintenance of the topological structure of ant–plant mutualistic networks.
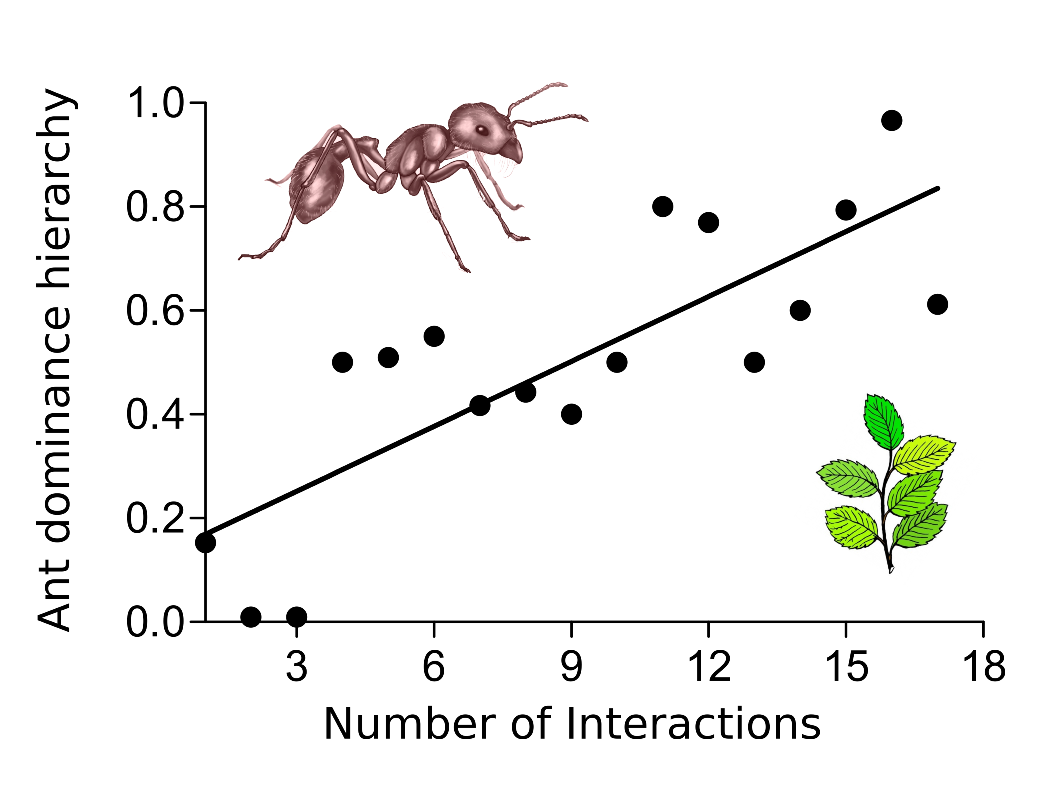
Figure 2. Correlation between the number of interactions and the dominance hierarchy in an ant community associated to EFN-bearing plants in a tropical coastal environment of the Gulf of Mexico (Adapted from Dáttilo et al. 2014b).
References
Dáttilo, W.; Guimarães, P.R.; Izzo, T.J. (2013). Spatial structure of ant-plant mutualistic networks. Oikos 122: 1643-1648. http://dx.doi.org/10.1111/j.1600-0706.2013.00562.x
Dáttilo, W.; Marquitti, F.M.D.; Guimarães, P.R.; Izzo, T.J. (2014a). The structure of ant-plant ecological networks: is abundance enough?. Ecology 95: 475-485. http://dx.doi.org/10.1890/12-1647.1
Dáttilo, W.; Díaz-Castelazo, C.; Rico-Gray, V. (2014b). Ant dominance hierarchy determines the nested pattern in ant-plant networks. Biological Journal of the Linnean Society 113: 405-414. http://dx.doi.org/10.1111/bij.12350
The authors
Wesley Dáttilo1; Thiago Junqueira Izzo2; Víctor Rico-Gray3
1PhD. Student in Animal Behavior at Universidad Veracruzana, Mexico – wdattilo@hotmail.com / www.wdattilo.bio.br
2Professor at Universidade Federal de Mato Grosso, Brazil – izzothiago@gmail.com
3Researcher at Universidad Veracruzana, Mexico – vrico@uv.mx
